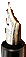

The evidence of carcass butchery by humans is old. Cut-marked bones found both in Gona and in Middle Awash (Ethiopia) show that systematic butchery might have begun around 2.5 million years ago. The small sample of cut-marked bone does not allow complete interpretation of the butchery practices of hominids at that time, but it clearly shows that they were defleshing (abundant number of cut marks on mid-shafts of upper and intermediate limb bones), dismembering, removing tongue, and even (at least, occasionally) eviscerating (Figure 1). Archaeologists still do not know if the sites where these cut-marked bones were found are kill-butchering sites or other type of sites where the remains were transported from the loci of carcass obtainment.
By 1.8 million years ago, a few sites in Kenya and Tanzania have evidence that hominids were processing carcasses in different types of sites. Sites where animals were initially obtained and butchered are not documented as well as those where those animals were repeatedly transported from loci of obtainment. The whole range of butchering practices is documented: defleshing, dismembering, eviscerating, tongue removal, and marrow processing.
Traditionally, assemblages consisting of stone tools and bones from a single animal or several animals of the same species have been interpreted as ‘butchering sites’. The oldest butchering sites thus documented were FLK North 6 at Olduvai (Tanzania) and FxJj3 (HAS) in Koobi Fora (Kenya), about 1.7-1.8 million years ago, where the remains of an elephant and hippopotamus, respectively, appeared spatially associated with stone tools. ‘Butchering sites’ are abundant during the Early and Middle Pleistocene (Table 1). Associations of stone tools and megafaunal remains have been widely documented in Olorgesailie and Koobi Fora (Kenya), Fejej, Gadeb and Middle Awash (Ethiopia), and Dandero (Eritrea). However, natural processes may also account for the spatial associations of lithic and bone remains. Most of these sites appear in alluvial contexts where natural clusters of bones from hippo and elephant carcasses are frequent, given the preferential use of these habitats by those animals. The taphonomic analyses of archaeological ‘butchering sites’ from this time period have revealed remains

Figure 1 Electronic microscope (SEM) photo of several cut marks found in a 2.5-million-year-old bone from Gona (Ethiopia).
From different taxa found together in a single site (see Taphonomy). In addition, in most of these sites, not a single bone showed any modification created by human butchering activities. In several sites, such as FLK North 6, Mwanganda, or Karonga, it has been observed that artifacts underlie the elephant bones. This suggests a deposition of artifacts previous to (and independent of) the deposition of the elephants. A recent revision of FLK North 6 has shown that the lithics and bones in the assemblage are not functionally related and the spatial association is inadvertent. The author has also revised the Gadeb butchering sites and the depositional histories of bones and stone tools are different and independent. From a taphonomic point of view, none of these assemblages can be interpreted as solid evidence of butchering sites.
The only sustainable claims of butchering sites in Africa from this time period are from the elephant site at Olorgesailie (Kenya) (an exclusive accumulation of an elephant and lithic artifacts whose functional link is pending taphonomic confirmation) and Peninj (Tanzania). This is also evidenced later in the Herto sites (end of Middle Pleistocene) in Ethiopia, where hippo remains associated with stone tools bear cut marks. However, this reductionist view, focused on cut marks alone, prevents us from considering some archaeological sites where butchery of large fauna might have occurred. Modern observations of elephant and hippo carcass butchery show that a complete carcass may be butchered without leaving a single trace. The problem that archaeologists face is: how to differentiate these sites from others where the spatial association of lithic and bones is accidental? No answer to this question can be currently provided. This problem is exacerbated, for example, in European Middle Pleistocene open-air sites, where elephant remains are frequently mixed with bones from other animals. In most of these sites, the elephant bones have been interpreted as naturally deposited, given their fragmentary state. Following this assumption, the fact that large fauna remains may naturally appear within bone assemblages of other animals reinforces the idea that this could also be the case in sites where only megafaunal remains and artifacts exist. Both megafauna and hominids were very active in the same alluvial environments and spatial associations of lithic and bones may be completely accidental, as several sites have shown. In this case, the only evidence of butchery that is scientifically reliable is the presence of cut and percussion marks, and any physical signatures of hammerstone bone breakage, meaning that several potential butchering sites are likely archaeologically invisible.
Peninj preserves some 1.3-million-year-old butchering sites. One of them (ST4) is characterized by the
Confirmed taphonomic evidence refers to presence of cut or percussion marks.
Partial semi-articulated distribution of different anatomical units of an extinct giraffe, with spatially differentiated distribution of taxa, which resembles the distribution of bones in Hadza kill sites and might be indicative of the place having acted as a butchering spot (Figure 2). The spatial association of bones and stone tools would support this (given the presence of cut marks and percussion marks), as well as the low density of artifacts and the presence of clusters of articulated bones. Elements in the home bases of modern foragers tend to be more widely mixed and dispersed. Pseudoarticulation of elements in home bases is also marginal or nonexistent. Several anatomical units of at least three different animals have been accumulated and manipulated by hominids in a spatially restricted area of the base of the ST4 site. The evidence from the ST4 site supports that Lower Pleistocene hominids were processing carcasses and not just dismembering bones.
The virtual lack of any physical evidence of butchery of megafauna during the early Pleistocene contrasts with the presence of cut-marked bone at European Middle Pleistocene butchery sites. In Table 1, more than one out of every three sites from this period is assumed to represent megafaunal butchery episodes, have butchery traces in the form of cut marks. This supports the presence of butchering sites during this time, although none of them could be called ‘kill sites’, since the only documented hominid activity is butchery (in some cases, marginal butchery). The scarcity of artifacts at some of these sites, such as Torralba and Ambrona, is more suggestive of marginal butchery than complete butchery of elephant skeletons. The abundant presence of artifacts at other localities, such as the Boxgrove sites, is suggestive of more intensive butchery of large fauna (rhinoceros) and smaller animals (horse). Either way, access to these animals through scavenging cannot be differentiated from hunting. Therefore, the term ‘kill site’ applied to megafauna is only appropriate for Holocene sites, especially those occurring in historical times, where we have a more accurate record of how these animals were obtained.
The Torralba and Ambrona sites, formerly interpreted as mass kills of proboscideans, have been reanalyzed from a taphonomic point of view. Most of the elephant bone breakage is diagenetic; when there was green breakage, it was caused by carnivores. Very few remains bear traces of hominid butchery, and weathering stages of bones together with detailed microstratigraphic analysis of both sites show that carcass accumulation was due to long processes involving the deposition of single carcasses during long time intervals. Hominids seem to have played a very marginal role in the accumulations of bones at both sites, which are due to natural causes.
The only Middle Pleistocene site that could represent a mass kill site is La Cotte de Saint Brelade, in the island of Jersey close to Normandy, where a pile of bones of woolly mammoth and rhino, some of them bearing cut marks, were located at the bottom of a cliff. It has been interpreted as the result of Middle
Figure 2 Portion of the excavation of the ST4 butchery site at Peninj (Tanzania). Most bones belong to a pigmy giraffe that was butchered by hominids. Bones appeared on a dry riverbed. Most freshly broken bones occurred around a lava boulderthat seems to have acted as the anvil where those bones were broken. The giraffe bones bear cut marks and percussion marks as a result of butchery (redrawn from Dominguez-Rodrigo etal., 2007).
Pleistocene hominids having driven herds of these animals to the cliff from which they fell and died. Two distinctive bone piles found in layers 3 and 6 (isotopic stage 6) of 20 mammoths and 5 rhinos comprise virtually all the fauna of these levels. A smaller frequency of stone tools than the other levels where such bone piles do not exist suggests more episodic or brief occupation as expected at butchery sites. The rhino sample is mostly composed of infantile and juvenile individuals. Mammoths are represented by all age groups. Some of the mammoths show partial skeletons. Cut marks have been found on several bones of these animals. Cut marks have been observed on proximal ends of tusks (removal), on scapulas, and long bone shafts (defleshing). Given the presence of this mass of animals inside the ravine at the foot of a 70 m cliff, the previous characteristics support a mass death event (or more than one) better than isolated death events for each individual. Animals grazing upland plateau would have been driven along a headland to the edge of the cliff where they would have fallen. Something similar has been suggested for Le Solutre (France) toward the end of the Upper Pleistocene, where a bone pile of horses was excavated.
Bone piles containing individuals of the same species have also been documented in prehistoric North America (Table 1). Holocene kill sites in this geographic region are very abundant. Most of them are mass buffalo kill sites. In this case, carcasses of dozens of buffaloes have been unearthed at a single spot. Archaeologists working on these sites have differentiated between a ‘kill’ site and a ‘butchering’ site. Kill sites are identified in proximity of a natural or contrived trap; the bone bed consists of bones from a single species, showing a high degree of articulated bones, with a corresponding proportion of butchering tools; and bones accumulated are frequently unbroken. Butchering sites are spots with smaller density of remains, absence of those elements usually discarded at kill sites (e. g., skulls), a lower frequency of articulated elements, and lack of complete skeletons. However, some of these assertions are ambiguous, since they also occur in natural palaeontological associations. These mass kill sites have been interpreted as the result of humans driving herds of bison to natural traps where they are collectively hunted. Corral structures have been discovered at some of these sites (e. g., Ruby). Most frequently, these sites occur in natural traps like streams or arroyos (e. g., Hawken), narrow valleys or sand dunes (e. g., Casper), bottom of cliffs (e. g., Glenrock Buffalo Jump), karsts (e. g., Vore), etc. Some sites, like the latter, show a thick bone level representing a single season kill of hundreds of animals.
The most widespread type of zooarchaeological analysis at these sites is skeletal part representation. Other data such as cut mark frequency and distribution, bone breakage, and intervention of other nonhuman agents in bone modification are only marginally considered. By taking anatomical data into consideration, it can be observed that there is a large variation in element representation. This suggests that either the functionality of these sites was different among one another, taphonomic processes affecting site formation were also diverse, or that hunting techniques were more varied than historically documented.
Butchering sites of larger animals, such as mammoth, have also been documented in North America during the end of the Pleistocene (see Table 1). In most cases, remains belong to single individuals, which suggest that hunting techniques for mammoths and bison were different.
In general, compared to other types of sites (workshops, central places), butchering sites are not abundant in the archaeological record. Table 1 Summed up the most representative ‘kill’ or ‘butchering’ sites for 2.5 million years of archaeological record. It can be argued that butchering sites during the Middle Pleistocene can be taphonomically confirmed. In contrast, very few sites of this type can be documented for earlier periods. During the Upper Pleistocene and Holocene, butchering and kill sites are more abundant and can be more easily confirmed from a tapho-nomic point of view.
See also: Extinctions of Big Game; Taphonomy; Vertebrate Analysis.
 World History
World History









